Xavier Morin
Cell division and neurogenesis
Goal
To understand how neural stem cells choose between proliferation and differentiation during development of the nervous system.
Background
The size, organization, and function of the brain depend on when, where, and in which quantities neurons and glial cells are produced during development. These cells derive from a limited reservoir of neural stem cells, which lose their capacity to proliferate as they differentiate and become mature. Therefore the developing brain must maintain a balance between its ability to grow (proliferation) and the production of functional neural cells (differentiation) as it matures. This is achieved through a mechanism of asymmetric cell division (ACD), during which a neural stem cell divides to self-renew and to produce a differentiating progeny. The mechanisms controlling asymmetric neurogenic divisions, as opposed to symmetric proliferative divisions, are not yet understood. Studying this phenomenon is a crucial challenge, not only at the basic level, but also to better understand the origin of developmental neurodisorders such as microcephaly. These questions are at the heart of the research conducted in Xavier Morin’s laboratory.
Research highlights
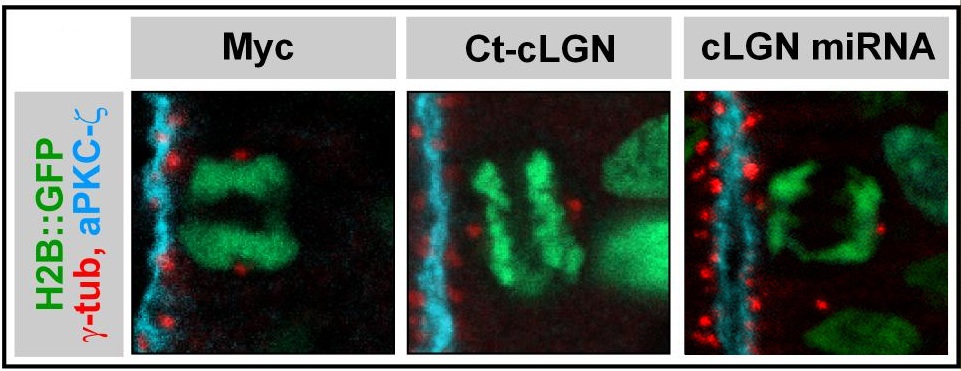
In many models, choices of ’asymmetrical’ identity between sister cells are correlated with the orientation of the axis of division of the mother cell in the tissue. This has led to the widely accepted idea that the orientation controls the fate of the daughter cells. This hypothesis has prompted us to explore the mechanisms that control the orientation of cell divisions, and we have thus helped to identify and characterize a molecular cascade largely conserved during evolution and used in both symmetrical and asymmetric divisions (Morin et al, 2007, Peyre et al, 2011, Saadaoui et al, 2014, 2017, di Pietro et al, 2017). However, the study of developmental defects associated with a disruption of the division axis of neural progenitors shows that it has little or no direct effect on the acquisition of daughter cell identity (stem cell or neuron). It seems that, at least in the neuroepithelium of the spinal cord, on which we carry out most of our work, the highly regulated planar orientation of the mitotic spindle mainly responds to the need to preserve the architecture of this tissue by maintaining the two daughter cells within the plane of the epithelial sheet after mitosis (Morin et al, 2007).
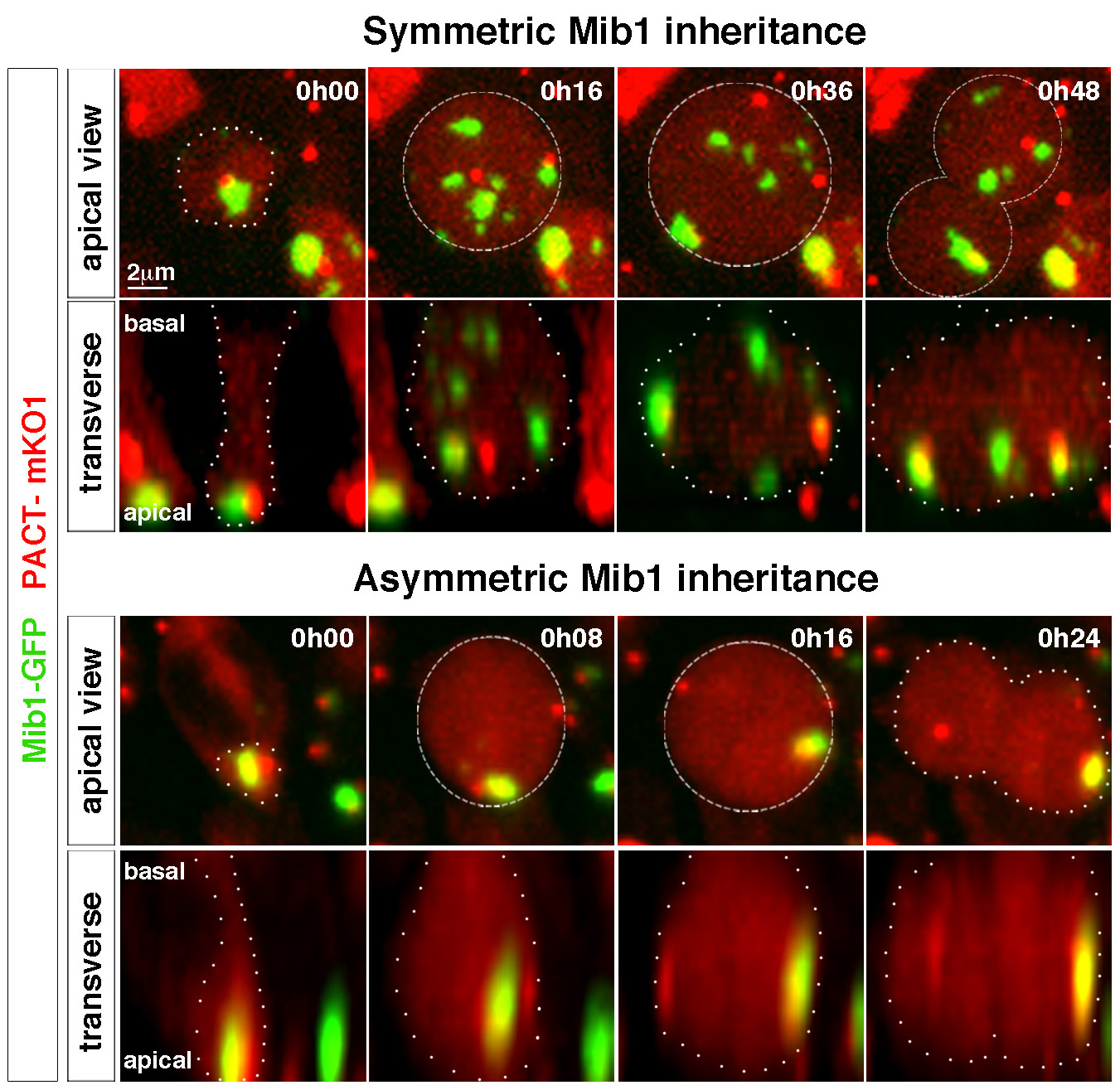
In parallel, we have started to explore other mechanisms that could control the asymmetry of division in neural stem cells. Thus, we recently observed the asymmetric distribution of a regulator of the Notch signaling pathway, the ubiquitin ligase Mindbomb (Mib1) in dividing neuroepithelial progenitors. Mib1, associated with components of the pericentriolar satellite (a poorly known organelle that “gravitates” around centrioles), is enriched near the "younger" centriole in interphase, which results in its location at a single pole of the mitotic spindle, and its inheritance after division by a single daughter cell which will become a neuron. By combining gain and loss of function experiments with real-time imaging of cell division and differentiation in the chicken neural tube, we have shown that this asymmetry of Mib1 distribution is instructive for sister cell identity. Even more remarkable, we have also shown that some of the progenitors "escape" this asymmetry thanks to the presence of an additional pool of Mib1 stored at the level of the Golgi apparatus in interphase, and sent to the daughter cell which does not receive the centrosomal pool of Mib1 (Tozer et al, 2017). In agreement with a progressive transition between the different modes of division, we observed at the tissue scale that the localization of Mib1 evolves between the early phase of proliferation and the later stages of neurogenesis.
Current Projects
These results are particularly intriguing and raise new questions. How is Mib1 recruited to the Golgi apparatus, and what are the molecular partners that regulate the Mib1 traffic between Golgi and centriolar satellites to control fate choices? A known role of Mib1 is to modify Dll1, a Notch ligand, at the cell membrane: therefore we think likely that Mib1 is essentially active in this location, while Golgi and centriolar satellites essentially fulfill a role of storage and routing. When and how does Mib1 transit between satellites and the plasma membrane? Answering all these questions requires high-resolution tracking of the sub-cellular localization of Mib1 over time, both during cell cycle progression and developmental time. It is also necessary to be able to purify Mib1 to identify its specific binding partners in different cellular compartments. We are developing the tools to respond to these questions.
At the same developmental stage in the neuroepithelium, symmetrical and asymmetrical divisions are simultaneously observed, but within the same cell clone, the two division modes follow one another and the transition between symmetrical and asymmetrical divisions seems irreversible, suggesting that the choice of division mode is not stochastic but highly controlled. Thus, beyond the question of the implementation of asymmetry for the acquisition of a different identity between daughter cells in neurogenic divisions, this implies that an upstream choice, under the control of specific genetic programs, determines the mode division of progenitors between symmetric (proliferative) and asymmetric (neurogenic). This question, which has been little discussed in "historical" models of asymmetric division based on invariant lineages in invertebrates, is at the heart of a new project developed in the team, which aims to identify these programs by studying the differences of transcriptional profile and organization of chromatin between progenitors in symmetric versus asymmetrical division mode.
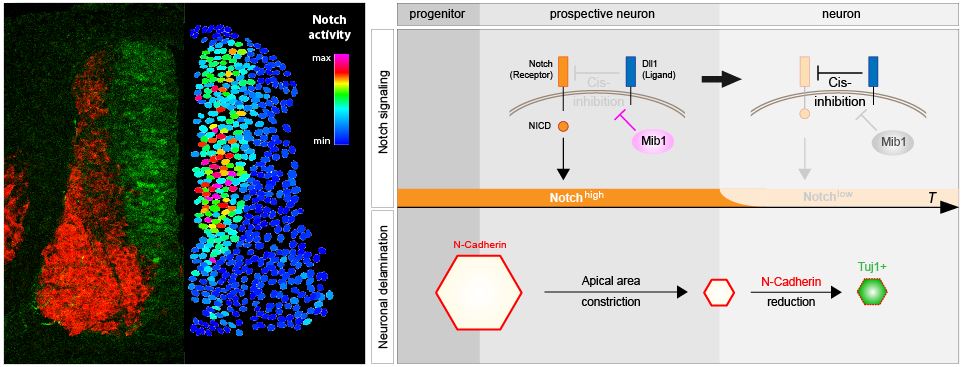
Baek C, Freem L, Goïame R, Sang H, Morin X, Tozer S. Mib1 prevents Notch Cis-inhibition to defer differentiation and preserve neuroepithelial integrity during neural delamination.
PLoS Biology 2018, 16(4):e2004162.
di Pietro F, Valon L, Li Y, Goïame R, Genovesio A, Morin X. An RNAi screen in a novel model of oriented divisions identifies the actin capping protein Z β as an essential regulator of spindle orientation.
Current Biology 2017, 27(16):2452–2464
Saadaoui M, Konno D, Loulier K, Goiame R, Jadhav V, Mapelli M, Matsuzaki F, Morin X. Loss of the canonical spindle orientation function in the Pins/LGN ortholog AGS3.
EMBO Reports 2017, 18(9):1509-1520
Tozer S, Baek C, Fischer E, Goïame R, Morin X. Differential routing of Mindbomb1 via centriolar satellites regulates asymmetric divisions of neural progenitors.
Neuron 2017, 93(3):542-551.
Saadaoui M, Machicoane M, di Pietro F, Etoc F, Echard A, Morin X. Dlg1 controls planar spindle orientation in the neuroepithelium through direct interaction with LGN.
The Journal of Cell Biology 2014, 206(6):707-717.
Loulier K, Barry R, Mahou P, Le Franc Y, Supatto W, Matho KS, Siohoi I, Fouquet S, Dupin E, Benosman R, Chedotal A, Beaurepaire E, Morin X, Livet J. Multiplex cell and lineage tracking with combinatorial labels.
Neuron 2014, 81(3):505-520.